LA CELULA Y SUS FUNCIONES.
ORGANIZACION DE LA CELULA:
Sus dos partes mas importantes son el núcleo y el citoplasma, separados entre si por una membrana nuclear, mientras que el citoplasma esta separado de los líquidos circundantes por una membrana celular también llamada membrana plasmática. Las diferentes sustancias que componen la célula se conocen como protoplasma. este esta compuesto por: agua, electrolitos, proteínas, líquidos e hidratos de carbono.

Agua. El principal medio líquido de la célula es el agua, que está presente en la mayoría de las células, excepto en los adipocitos, en una concentración del 70-85%.
Iones. Algunos de los iones importantes de la célula son el potasio, el magnesio, él fosfato, el sulfato, el bicarbonato y cantidades más pequeñas de sodio, cloruro y calcio. Los iones son los productos químicos inorgánicos de las reacciones celulares y son necesarios para el funcionamiento de algunos de los mecanismos de control celulares.
Proteínas. Después del agua, las sustancias más abundantes en la mayoría de las células son las proteínas, que normalmente constituyen entre el 10 y el 20% de la masa celular. Son de dos tipos, proteínas estructurales y proteínas funcionales. Las proteínas estructurales están presentes en la célula principalmente en forma de filamentos largos que son polímeros de muchas moléculas proteicas individuales. Las proteínas funcionales son un tipo de proteína totalmente diferente, compuesto habitualmente por combinaciones de pocas moléculas en un formato tubular-globular. Estas proteínas son principalmente las enzimas de la célula y, al contrario de las proteínas fibrilares, a menudo son móviles dentro del líquido celular. Además, muchas de ellas están adheridas a las estructuras membranosas dentro de la célula.
Lípidos. Los lípidos son varios tipos de sustancias que se agrupan porque tienen una propiedad común de ser solubles en disolventes grasos. Lípidos especialmente importantes son los fosfolípidos y el colesterol, que juntos suponen sólo el 2% de la masa total de la célula. Su importancia radica en que, al ser principalmente insolubles en agua, se usan para formar las barreras de la membrana celular y de la membrana intracelular que separan los distintos compartimientos celulares.

Hidratos de carbono. Los hidratos de carbono tienen escasas funciones estructurales en la célula, salvo porque forman parte de las moléculas glucoproteicas, pero sí tienen un papel muy importante en la nutrición celular. Los hidratos de carbono siempre están presentes en forma de glucosa disuelta en el líquido extracelular circundante, de forma que es fácilmente accesible a la célula.
Estructura física de la célula
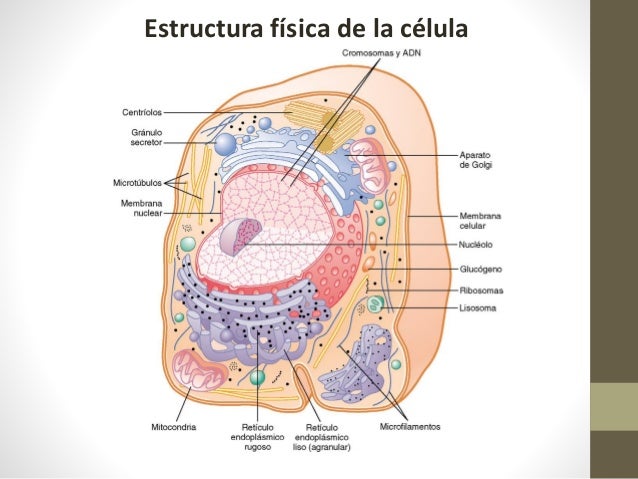
La célula no es una simple bolsa de líquido, enzimas y productos químicos, también contiene estructuras físicas muy organizadas que se denominan orgánulos intracelulares. Sin uno de los orgánulos, la mitocondria, más del 95% de la energía de la célula que se libera de los nutrientes desaparecería inmediatamente.
Estructuras membranosas de la célula
La mayoría de los orgánulos de la célula están cubiertos por membranas compuestas principalmente por lípidos y proteínas. Estas membranas son la membrana celular, la membrana nuclear, la membrana del retículo endoplásmico y las membranas de la mitocondria, los lisosomas y el aparato de Golgi. Los lípidos de las membranas proporcionan una barrera que impide el movimiento de agua y sustancias hidrosolubles desde un compartimiento celular a otro, porque el agua no es soluble en lípidos.
Membrana celular
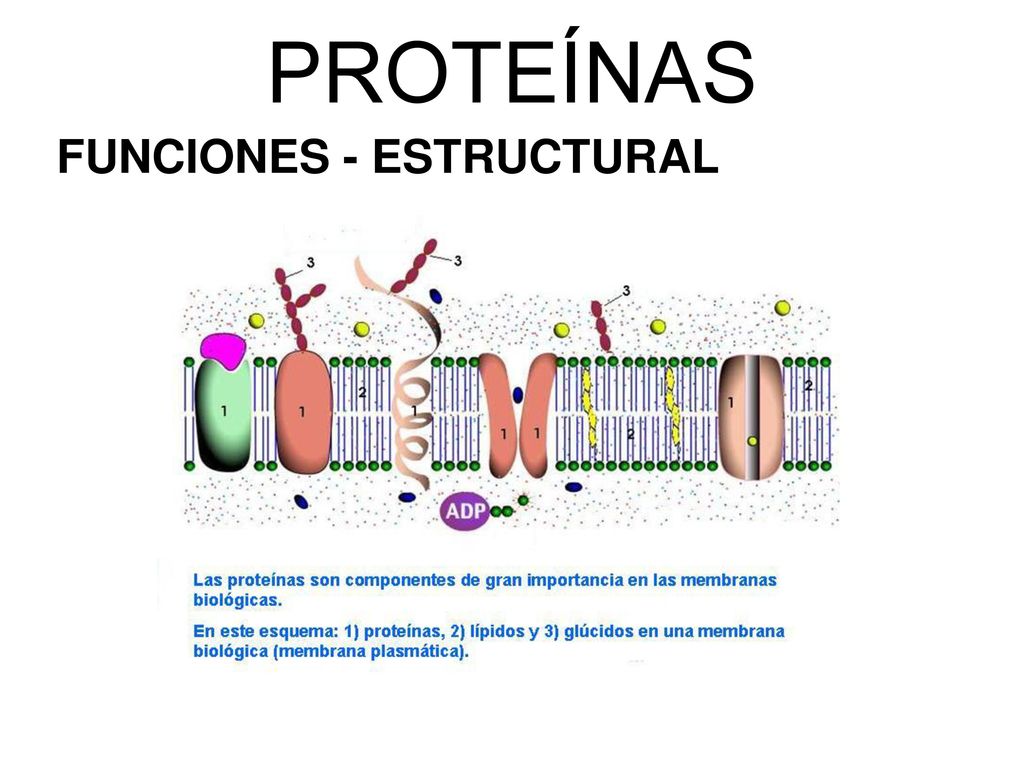
La membrana celular (también denominada membrana plasmática), que cubre la célula, es una estructura elástica, fina y flexible que tiene un grosor de tan sólo 7,5 a 10 nm. Está formada casi totalmente por proteínas y lípidos, con una composición aproximada de un 55% de proteínas, un 25% de fosfolípidos, un 13% de colesterol, un 4% de otros lípidos y un 3% de hidratos de carbono.

Su estructura básica consiste en una bicapa lipídica, una película fina de doble capa de lípidos, cada una de las cuales contiene una sola molécula de grosor y rodea de forma continua toda la superficie celular. En esta película lipídica se encuentran intercaladas grandes moléculas proteicas globulares. La bicapa lipídica básica está formada por moléculas de fosfolípidos. Un extremo de cada molécula de fosfolípido es soluble en agua, es decir, es hidrófilo, mientras que el otro es soluble sólo en grasas, es decir, es hidrófobo. El extremo fosfato del fosfolípido es hidrófilo y la porción del ácido graso es hidrófoba. La capa lipídica de la zona media de la membrana es impermeable a las sustancias hidrosolubles habituales, como iones, glucosa y urea. Por el contrario, las sustancias hidrosolubles, como oxígeno, dióxido de carbono y alcohol, pueden penetrar en esta porción de la membrana con facilidad. El colesterol también controla gran parte de la fluidez de la membrana.

Proteínas de la membrana celular integrales y periféricas.
Son proteínas de membrana, glucoproteínas en su mayoría. Existen dos tipos de proteínas de membrana celular: proteínas integrales que hacen protrusión por toda la membrana y proteínas periféricas que se unen sólo a una superficie de la membrana y que no penetran en todo su espesor. Muchas de las proteínas integrales componen canales estructurales (o poros) a través de los cuales las moléculas de agua y las sustancias hidrosolubles, especialmente los iones, pueden difundir entre los líquidos extracelular e intracelular. Estos canales de proteínas también tienen propiedades selectivas que permiten la difusión preferente de algunas sustancias con respecto a las demás. Otras proteínas integrales actúan como proteínas transportadoras de sustancias que, de otro modo, no podrían penetrar en la bicapa lipídica. Las proteínas integrales de la membrana pueden actuar también como receptores de los productos químicos hidrosolubles, como las hormonas peptídicas, que no penetran fácilmente en la membrana celular. Las moléculas proteicas periféricas se unen con frecuencia a las proteínas integrales, de forma que las proteínas periféricas funcionan casi totalmente como enzimas o como controladores del transporte de sustancias a través de los «poros» de la membrana celular

Hidratos de carbono de la membrana: «glucocáliz» celular. Los hidratos de carbono de la membrana se presentan casi invariablemente combinados con proteínas o lípidos en forma de glucoproteínas o glucolípidos. De hecho, la mayoría de las proteínas integrales son glucoproteínas y aproximadamente la décima parte de las moléculas lipídicas de la membrana son glucolípidos. Hay muchos otros compuestos de hidratos de carbono, que se denominan proteoglicanos y son principalmente hidratos de carbono unidos a núcleos de proteínas pequeñas, que también se unen laxamente a la superficie externa de la pared celular, es decir, toda la superficie externa de la célula a menudo contiene un recubrimiento débil de hidratos de carbono que se conoce como glucocáliz. Las estructuras de hidratos de carbono unidas a la superficie exterior de la célula tienen varias funciones importantes: 1) muchas de ellas tienen una carga eléctrica negativa que proporciona a la mayoría de las células una carga negativa a toda la superficie que repele a otros objetos negativos; 2) el glucocáliz de algunas células se une al glucocáliz de otras, con lo que une las células entre sí; 3) muchos de los hidratos de carbono actúan como componentes del receptor para la unión de hormonas, como la insulina; cuando se unen, esta combinación activa las proteínas internas unidas que, a su vez, activan una cascada de enzimas intracelulares, y 4) algunas estructuras de hidratos de carbono participan en reacciones inmunitarias.
Citoplasma y sus orgánulos
El citoplasma está lleno de partículas diminutas y grandes y orgánulos dispersos. La porción de líquido del citoplasma en el que se dispersan las partículas se denomina citosol y contiene principalmente proteínas, electrólitos y glucosa disueltos. En el citoplasma se encuentran dispersos glóbulos de grasa neutra, gránulos de glucógeno, ribosomas, vesículas secretoras y cinco orgánulos especialmente importantes: el retículo endoplásmico, el aparato de Golgi, las mitocondrias, los lisosomas y los peroxisomas.

Retículo endoplásmico
La superficie total de esta estructura en algunas células, como los hepatocitos, por ejemplo, puede ser hasta 30 o 40 veces la superficie de la membrana celular. El espacio que queda dentro de los túbulos y vesículas está lleno de una matriz endoplásmica, un medio acuoso que es distinto del líquido del citosol que hay fuera del retículo endoplásmico. Las microfotografías electrónicas demuestran que el espacio que queda dentro del retículo endoplásmico está conectado con el espacio que hay entre las dos superficies de la membrana nuclear. Las sustancias que se forman en algunas partes de la célula entran en el espacio del retículo endoplásmico y después son conducidas a otras partes de la célula.
Ribosomas y retículo endoplásmico rugoso. Unida a la superficie exterior de muchas partes del retículo endoplásmico nos encontramos una gran cantidad de partículas granulares diminutas que se conocen como ribosomas. Cuando están presentes, el retículo se denomina retículo endoplásmico rugoso. Los ribosomas están formados por una mezcla de ARN y proteínas y su función consiste en sintetizar nuevas moléculas proteicas en la célula.
Retículo endoplásmico agranular. Parte del retículo endoplásmico no tiene ribosomas, es lo que se conoce como retículo endoplásmico agranular, o liso. Este retículo agranular actúa en la síntesis de sustancias lipídicas y en otros procesos de las células que son promovidos por las enzimas intrarreticulares.

Aparato de Golgi.
Está íntimamente relacionado con el retículo endoplásmico. Tiene unas membranas similares a las del retículo endoplásmico agranular y está formado habitualmente por cuatro o más capas apiladas de vesículas cerradas, finas y planas, que se alinean cerca de uno de los lados del núcleo. Este aparato es prominente en las células secretoras, donde se localiza en el lado de la célula a partir del cual se extruirán las sustancias secretoras. El aparato de Golgi funciona asociado al retículo endoplásmico.

Las sustancias atrapadas en las vesículas del RE se transportan desde el retículo endoplásmico hacia el aparato de Golgi. Las sustancias transportadas se procesan después en el aparato de Golgi para formar lisosomas, vesículas secretoras y otros componentes citoplasmáticos.
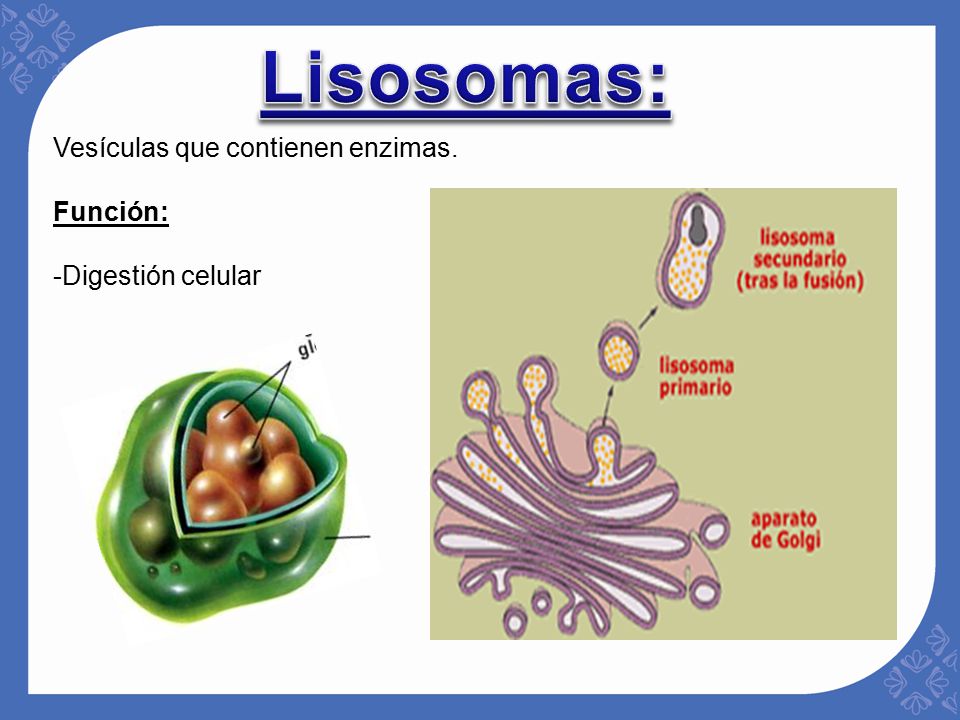
Lisosomas.
Son orgánulos vesiculares que se forman por la rotura del aparato de Golgi y después se dispersan por todo el citoplasma. Los lisosomas constituyen el aparato digestivo intracelular que permite que la célula digiera: 1) las estructuras celulares dañadas; 2) las partículas de alimento que ha ingerido, y 3) las sustancias no deseadas, como las bacterias. El lisosoma es muy distinto en los distintos tipos celulares, pero habitualmente tiene un diámetro de 250 a 750 nm. Está rodeado por una membrana bicapa lipídica típica y está llena con grandes cantidades de gránulos pequeños, de 5 a 8 nm de diámetro, que son agregados de proteínas que contienen hasta 40 tipos diferentes de enzimas (digestivas) de tipo hidrolasa. Lo normal es que la membrana que rodea los lisosomas impida que las enzimas hidrolíticas encerradas en ellos entre en contacto con otras sustancias de la célula y, por tanto, previene sus acciones digestivas.
Peroxisomas
Los peroxisomas son similares físicamente a los lisosomas, pero son distintos en dos aspectos importantes. En primer lugar, se cree que están formados por autorreplicación (o, quizás, haciendo protrusión desde el retículo endoplásmico liso) en lugar de proceder del aparato de Golgi. En segundo lugar, contienen oxidasas en lugar de hidrolasas. . El peróxido de hidrógeno es una sustancia muy oxidante que se utiliza junto a una catalasa, otra enzima oxidasa que se encuentra en grandes cantidades en los peroxisomas para oxidar muchas sustancias que, de lo contrario, serían venenosas para la célula.
Vesículas secretoras
Una de las funciones importantes de muchas células es la secreción de sustancias químicas especiales. Casi todas las sustancias secretoras se forman en el sistema retículo endo- plásmico-aparato de Golgi y después de liberan desde el aparato de Golgi hacia el citoplasma en forma de vesículas de almacenamiento que se conocen como vesículas secretoras o gránalos secretores.

Mitocondrias. Se conocen como los «centros neurálgicos» de la célula. Sin ellas, las células no serían capaces de extraer energía suficiente de los nutrientes y, en esencia, cesarían todas las funciones celulares. Las mitocondrias se encuentran en todas las zonas del citoplasma de la célula, pero su número total en cada célula varía de menos de cien hasta varios miles, dependiendo de la cantidad de energía que requiere la célula. Algunas miden sólo algunos cientos de nanómetros de diámetro y adoptan forma globular, mientras que otras son alargadas, miden hasta 1 fxm de diámetro y 7 (Jim de longitud; un tercer tipo tiene una estructura ramificada y filamentosa. La estructura básica de la mitocondria, está compuesta principalmente por dos membranas de bicapa lipídica-proteínas: una membrana externa y una membrana interna. La cavidad interna de la mitocondria está llena con una matriz que contiene grandes cantidades de enzimas disueltas que son necesarias para extraer la energía de los nutrientes.
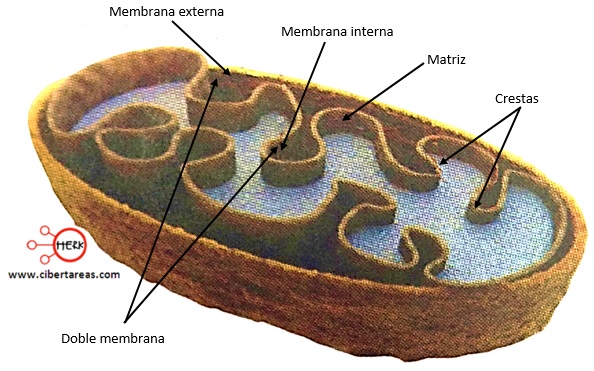
Las mitocondrias se reproducen por sí mismas, lo que significa que una mitocondria puede formar una segunda, una tercera, etc., siempre que la célula necesite cantidades mayores de ATP.
Citoesqueleto celular: estructuras filamentosas y tubulares. Todas las células usan un tipo especial de filamento rígido formado por polímeros de tubulina para construir estructuras tubulares fuertes, los microtúbulos. Una de las funciones principales de los microtúbulos es actuar como citoesqueleto, proporcionando estructuras físicas rígidas para determinadas partes de las células.
Núcleo
El núcleo es el centro de control de la célula. Brevemente, contiene grandes cantidades de ADN, es decir, los genes, que son los que determinan las características de las proteínas celulares, como las proteínas estructurales, y también las enzimas intracelulares que controlan las actividades citoplasmáticas y nucleares. Los genes también controlan y promueven la reproducción de la célula. Los genes se reproducen primero para obtener dos juegos idénticos de genes y después se divide la célula utilizando un proceso especial, que se conoce como mitosis, para formar dos células hijas, cada una de las cuales recibe uno de los dos juegos de genes de ADN.

Membrana nuclear
La membrana nuclear, también conocida como cubierta nuclear, consiste realmente en dos membranas bicapa separadas, una dentro de la otra. Varios miles de poros nucleares atraviesan la membrana nuclear. En los bordes de estos poros hay unidos grandes complejos de moléculas proteicas, de forma que la zona central de cada poro mide sólo unos 9nm en diámetro, tamaño suficientemente grande como para permitir que moléculas de un peso molecular de hasta 44.000 la atraviesen con una facilidad razonable.
Nucléolos y formación de ribosomas
Los núcleos de la mayoría de las células contienen una o más estructuras que se tiñen intensamente y se denominan nucléolos. Estos nucléolos, a diferencia de la mayoría de los orgánu- los que vamos a comentar, no tienen una membrana limitante, sino que consisten en una acumulación simple de grandes cantidades de ARN y proteínas de los tipos encontrados en los ribosomas. El nucléolo aumenta de tamaño considerablemente cuando la célula está sintetizando proteínas activamente. La formación de los nucléolos (y de los ribosomas del citoplasma fuera del núcleo) comienza en el núcleo.

Comparación entre la célula animal y las formas de vida precelulares. La célula es un organismo complicado que ha necesitado muchos cientos de millones de años para desarrollarse después de que apareciera la primera forma de vida, un organismo similar a los virus de nuestros días, sobre la tierra.
El componente vital esencial de los virus pequeños es un ácido nucleico embebido en un recubrimiento proteico. Este ácido nucleico está formado por los mismos componentes del ácido nucleico de base (ADN o ARN) que se encuentran en las células de mamíferos y es capaz de reproducirse a sí mismo en las condiciones apropiadas, es decir, que el virus propaga su linaje de generación en generación y, por tanto, es una estructura viva igual que lo son la célula y el ser humano. En etapas aún más avanzadas de la vida, en particular en las etapas de rickettsias y bacterias, se desarrollaron orgánulos dentro del organismo que representaban estructuras físicas de agregados químicos que realizan funciones de una forma más eficiente que la lograda por los productos químicos dispersos en la matriz líquida. Por último, en la célula nucleada se desarrollaron orgánulos aún más complejos, el más importante de los cuales es el propio núcleo. El núcleo distingue este tipo de célula de todas las demás formas de vida, proporciona un centro de control para todas las actividades celulares y también logra la reproducción exacta de una generación tras otra de células nuevas, teniendo cada nueva célula casi exactamente la misma estructura que su progenitor.
Sistemas funcionales de la célula.
Ingestión por la célula: endocitosis
Si una célula va a vivir, crecer y reproducirse, debe obtener nutrientes y otras sustancias de los líquidos circundantes. La mayoría de estas sustancias atraviesan la membrana celular por difusión y transporte activo. La difusión implica el movimiento simple a través de la membrana, provocado por el movimiento aleatorio de las moléculas de la sustancia; las sustancias se desplazan a través de los poros de la membrana celular o, en el caso de las sustancias liposolubles, a través de la matriz lipídica de la membrana. El transporte activo implica el transporte real de una sustancia a través de la membrana mediante una estructura física de carácter proteico que penetra en todo el espesor de la membrana. Las partículas muy grandes entran en la célula mediante una función especializada de la membrana celular que se denomina endocitosis. Las formas principales de endocitosis son la pinocitosis y la fagocitosis. La pinocitosis se refiere a la ingestión de partículas diminutas que forman vesículas de líquido extracelular y partículas dentro del citoplasma celular. La fagocitosis se refiere a la ingestión de partículas grandes, como bacterias, células enteras o porciones de tejido degenerado.
Pinocitosis. La pinocitosis se produce continuamente en las membranas celulares de la mayoría de las células, pero es especialmente rápida en algunas de ellas. La pinocitosis es el único medio por el cual las principales macromoléculas grandes, como la mayoría de las moléculas proteicas, pueden entrar en las células. De hecho, la velocidad con que se forman las vesículas de pinocitosis suele aumentar cuando estas macromoléculas se unen a la membrana celular. Este proceso requiere el aporte de energía desde el interior de la célula, que es suministrada por el ATP, un producto de alta energía. Además, requiere la presencia del ion calcio en el líquido extracelular, que probablemente reaccionará con los filamentos de proteína contráctil que hay por debajo de las hendiduras revestidas para proporcionar la fuerza que se necesita para que se produzca la separación de las vesículas lejos de la membrana celular.
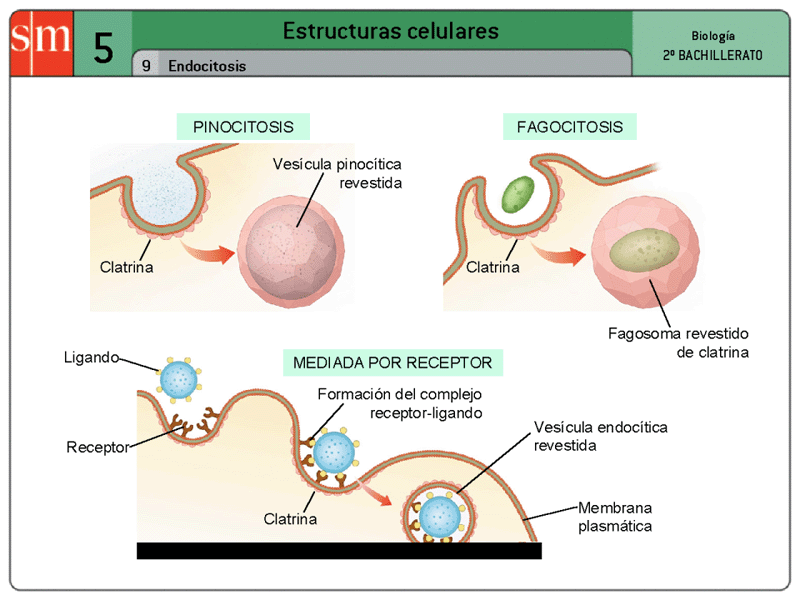
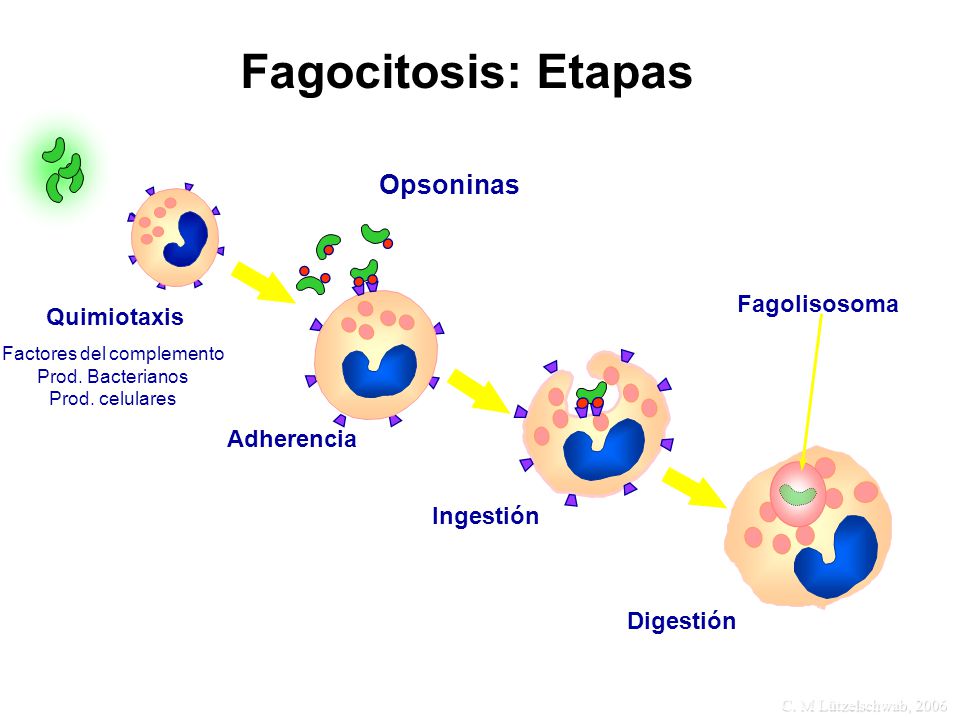
Fagocitosis. La fagocitosis se produce del mismo modo que la pinocitosis, en grandes rasgos, excepto porque implica la participación de partículas grandes y no moléculas. Sólo algunas células tienen la capacidad de realizar la fagocitosis, principalmente los macrófagos tisulares y algunos de los leucocitos sanguíneos. La fagocitosis se inicia cuando una partícula, como una bacteria, una célula muerta o un resto de tejido, se une a los receptores de la superficie de los fagocitos. En el caso de las bacterias, cada una de ellas ya suele estar unida a un anticuerpo específico frente a ese organismo y es ese anticuerpo el que se une a los receptores de fagocitosis, arrastrando consigo a la bacteria.
Regresión de los tejidos y autólisis de las células. Los tejidos del organismo a menudo regresan a un tamaño más pequeño. Los lisosomas son responsables de gran parte de esta regresión. Se desconoce el mecanismo por el que la falta de actividad de un tejido hace que los lisosomas aumenten su actividad. El daño de una célula causado por el calor, el frío, un traumatismo, productos químicos o cualquier otro factor induce la rotura de los lisosomas. Las hidrolasas liberadas comienzan inmediatamente a digerir las sustancias orgánicas circundantes. Si el daño es pequeño, sólo se eliminará una porción de la célula, que después se repararía. Si el daño es importante se digiere toda la célula, lo que se denomina autólisis. Los lisosomas también contienen sustancias bactericidas que pueden matar a las bacterias fagocitadas antes de que puedan provocar daños a la célula. Estas sustancias son: 1) la lisozima, que disuelve la membrana celular bacteriana; 2) la lisoferrina, que se une al hierro y a otras sustancias antes de que puedan promover el crecimiento bacteriano, y 3) un medio ácido, con un pH en torno a 5, que activa las hidrolasas e inactiva los sistemas metabólicos bacterianos.
Las proteínas se forman en el retículo endoplásmico rugoso. La porción granular del retículo endoplásmico se caracteriza por un gran número de ribosomas unidos a las superficies externas de la membrana del retículo endoplásmico.
Síntesis de lípidos en el retículo endoplásmico liso. El retículo endoplásmico también sintetiza lípidos, especialmente fosfolípidos y colesterol, que se incorporan rápidamente a la bicapa lipídica del propio retículo endoplásmico provocando que su crecimiento sea aún mayor. Esta reacción tiene lugar principalmente en la porción lisa del retículo endoplásmico.

Otras funciones del retículo endoplásmico. Otras funciones significativas del retículo endoplásmico, en especial del retículo liso, son las siguientes:
- Proporciona las enzimas que controlan la escisión del glucógeno cuando se tiene que usar el glucógeno para energía.
- Proporciona una cantidad inmensa de enzimas que son capaces de detoxificar las sustancias, como los fármacos, que podrían dañar la célula. Consigue la detoxificación por coagulación, oxidación, hidrólisis, conjugación con ácido glucurónico y de otras formas.
Funciones de síntesis del aparato de Golgi. Aunque una función importante del aparato de Golgi consiste en procesar todavía más las sustancias que ya se han formado en el retículo endoplásmico, también tiene la capacidad de sintetizar ciertos hidratos de carbono que no se pueden formar en el retículo endoplásmico, lo que es especialmente cierto para la formación de los grandes polímeros de sacáridos que se unen a cantidades pequeñas de proteínas; algunos ejemplos importantes son el ácido hialurónico y la condroitina sulfato. Algunas de las muchas funciones del ácido hialurónico y de la condroitina sulfato en el organismo son las siguientes: 1) son los principales componentes de los proteoglucanos segregados en el moco y en otras secreciones glandulares; 2) son los componentes principales de la sustancia fundamental que está fuera de las células en los espacios intersticiales, actuando como rellenos entre las fibras de colágeno y las células; 3) son los componentes principales de la matriz orgánica en el cartílago y en el hueso, y 4) son importantes en numerosas actividades celulares como la migración y la proliferación.
Tipos de vesículas formadas por el aparato de Golgi: vesículas secretoras y lisosomas. En una célula muy secretora las vesículas formadas por el aparato de Golgi son principalmente vesículas secretoras que contienen proteínas que se deben segregar a través de la superficie de la membrana celular. Estas vesículas secretoras difunden primero hacia la membrana celular, después se fusionan con ella y vacían sus sustancias hacia el exterior por el mecanismo denominado exocitosis. La exocitosis, en la mayoría de los casos, se estimula por la entrada de iones calcio en la célula; los iones calcio interaccionan con la membrana vesicular de alguna forma que no comprendemos y provocan su fusión con la membrana celular, seguida por exocitosis, es decir, la apertura de la superficie externa de la membrana y extrusión de su contenido fuera de la célula. No obstante, algunas vesículas están destinadas al uso intracelular.

Uso de vesículas intracelulares para reponer las membranas celulares. Algunas de las vesículas intracelulares que se forman en el aparato de Golgi se fusionan con la membrana celular o con las membranas de estructuras intracelulares, como la mitocondria e incluso el retículo endoplásmico, lo que aumenta la superficie de estas membranas y repone las membranas a medida que se van utilizando. Por ejemplo, la membrana celular pierde gran parte de su sustancia cada vez que forma una vesícula fagocítica o pinocítica y las membranas vesiculadas del aparato de Golgi reponen continuamente la membrana celular. En resumen, el sistema de membrana del retículo endoplásmico y el aparato de Golgi representa un órgano de un metabolismo intenso que es capaz de formar nuevas estructuras intracelulares, así como sustancias secretoras que se van a extruir de la célula.
Características funcionales del ATP.
El ATP es un nucleótido compuesto por: 1) la base nitrogenada adenina; 2) el azúcar pentosa ribosa, y 3) tres radicales fosfato. Los dos últimos radicales fosfato están conectados con el resto de la molécula mediante los denominados enlaces de fosfato de alta energía, que están representados en la fórmula representados por el símbolo ~. En las condiciones físicas y químicas del organismo cada uno de esos enlaces de alta energía contiene aproximadamente 12.000 calorías de energía por mol de ATP, cifra muchas veces mayor que la energía almacenada en un enlace químico medio, dando lugar al término enlace de alta energía. Cuando el ATP libera su energía se separa un radical de ácido fosfórico y se forma difosfato de adenosina (ADP). La energía liberada se usa para dar energía muchas de las demás funciones celulares, como la síntesis de sustancias y la contracción muscular. Para reconstituir el ATP celular conforme se gasta, la energía derivada de los nutrientes celulares hace que el ADP y el ácido fosfórico se recombinen para formar una nueva molécula de ATP y todo el proceso se repite una y otra vez. Por este motivo, el ATP se conoce como la moneda energética de la célula porque se puede gastar y recomponer continuamente, con un ciclo metabólico de sólo unos minutos.


Usos de ATP para las funciones celulares. La energía del ATP se usa para promover tres categorías principales de funciones celulares: 1) transporte de sustancias a través de múltiples membranas en la célula; 2) síntesis de compuestos químicos a través de la célula, y 3) trabajo mecánico. Estos usos del ATP se ilustran mediante los ejemplos de la figura 2-15: 1) para suministrar energía para el transporte de sodio a través de la membrana celular; 2) para favorecer la síntesis proteica en los ribosomas, y 3) para suministrar la energía necesaria durante la contracción muscular. Además del transporte de sodio en la membrana, la energía del ATP es necesaria para el transporte a través de la membrana de iones potasio, calcio, magnesio, fosfato, cloruro, urato, hidrógeno y muchos otros iones y varias sustancias orgánicas. El transporte en la membrana es tan importante para las funciones de la célula que algunas, como las del túbulo renal, consumen hasta el 80% del ATP que forman sólo para este propósito.

LOCOMOCION DE LA CELULA
Con mucho, el tipo más importante de movimiento que se produce en el organismo es el de los miocitos en el músculo esquelético, cardíaco y liso, que constituye casi el 50% de toda la masa del organismo. En otras células se producen otros tipos de movimiento, el amebiano y el ciliar.
Movimiento amebiano
El movimiento amebiano es el movimiento de toda la célula en relación con su entorno, como el movimiento de los leucocitos a través de los tejidos. Recibe su nombre por el movimiento de las amebas, que es de este tipo. El movimiento amebiano comienza con la protrusión de un seudópodo desde un extremo de la célula. Este seudópodo se proyecta a distancia, lejos de la célula, y se asegura parcialmente en una zona nueva. Después, tira del resto de la célula hacia él.

Mecanismo de locomoción amebiana.
Básicamente, es consecuencia de la formación continua de una membrana celular nueva en el extremo director del seudópodo y la absorción continua de la membrana en las porciones media y posterior de la célula.
Tipos de células que muestran movimiento amebiano. Las células más frecuentes que muestran movimiento amebiano en el cuerpo humano son los leucocitos cuando salen de la sangre hacia los tejidos para formar macrófagos tisulares. Otros tipos de células también pueden moverse con un movimiento amebiano en determinadas circunstancias. Por ejemplo, los fibroblastos se mueven hacia una zona dañada para reparar el daño e incluso las células germinales de la piel que, aunque normalmente son células totalmente sésiles, se desplazan hacia la zona de un corte para reparar el desgarro.
Control del movimiento amebiano: quimiotaxia. El iniciador más importante del movimiento amebiano es la quimiotaxia, proceso que se produce como consecuencia de la aparición de determinadas sustancias en el tejido. Cualquier sustancia que provoque la quimiotaxia se conoce como sustancia quimiotáctica y la mayoría de las células que utilizan movimientos amebianos se desplazan hacia el origen de la sustancia quimiotáctica, es decir, desde una zona de concentración más baja a otra de concentración más alta, es decir, una quimiotaxia positiva, mientras que otras se alejan del origen, o quimiotaxia negativa.
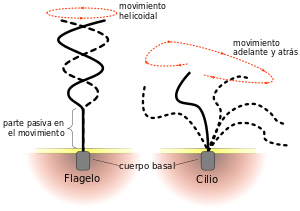
Cilios y movimientos ciliares
Un segundo tipo de movimiento celular, el movimiento ciliar, es un movimiento a modo de látigo de los cilios que se encuentran en la superficie de las células. Este movimiento existe sólo en dos lugares del cuerpo humano: en la superficie de las vías respiratorias y en la superficie interna de las trompas uterinas (trompas de Falopio) del aparato reproductor. Un cilio tiene el aspecto de un pelo recto o curvo con punta afilada que se proyecta 2-4 |xm desde la superficie de la célula. A menudo, muchos cilios se proyectan desde una sola célula, por ejemplo, hay hasta 200 cilios en la superficie de cada célula epitelial dentro de las vías aéreas. Cada cilio es una excrecencia de una estructura que se apoya inmediatamente por debajo de la membrana celular, el cuerpo basal del cilio. El flagelo de un espermatozoide es similar a un cilio; de hecho, tiene el mismo tipo de estructura y el mismo tipo de mecanismo contráctil. Sin embargo, este flagelo es mucho más largo y se desplaza con ondas de tipo cuasi-sinusoidal en lugar de movimientos de tipo látigo.